
Solanum stoloniferum

| Common Names | Fendler’s potato |
| Code | sto |
| Synonyms | S. ajuscoense, S. antopoviczii, S. boreale, S. candelarianum, S. fendleri, S. longipedicellatum, S. malinchense, S. neoantopoviczii, S. papita, S. polytrichon, S. tlaxcalense, S. tuberosum var. boreale, S. wightianum |
| Clade | Mixed |
| Series | Longipedicellata |
| Ploidy | Tetraploid (4x) |
| EBN | 2 |
| Tuberization Photoperiod | Unknown |
| Self-compatibility | Yes |
| Nuclear Genome | AB |
| Cytoplasmic Genome | W, D |
| Citation | Von Schlechtendal: Linnaea 8: 255. 1833. |
Description
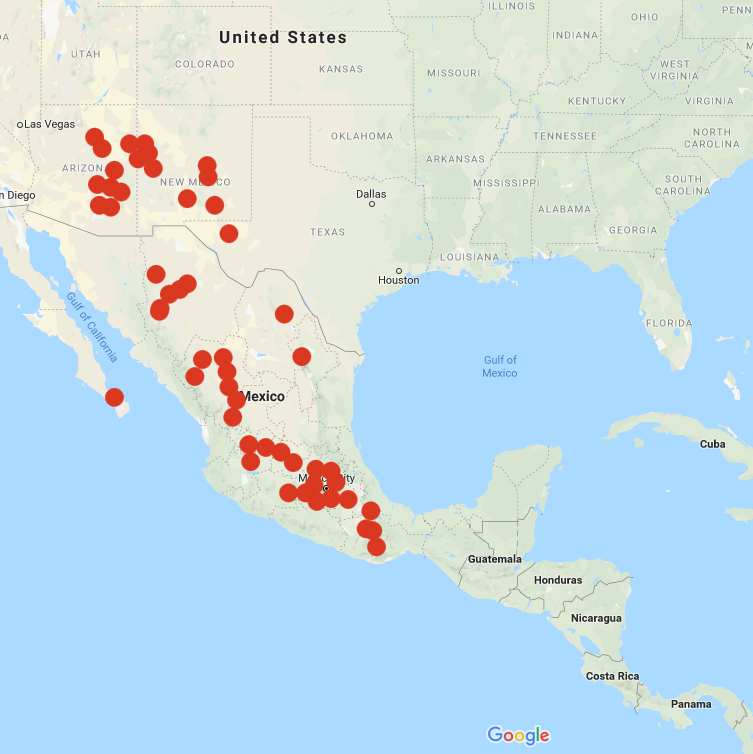
Solanum stoloniferum is a widespread and highly variable species, ranging in height from about 7 to 24 inches, ranging from the southwest US to southern Mexico. In the United States, it is found primarily in New Mexico and Arizona and reaches slightly into southwest Texas (Spooner 2004). It is one of only two wild potato species found in the United States and the only wild potato species found in Baja California.
The specific epithet, stoloniferum, means “bearing stolons,” which one might consider a rather obvious description for a potato. It is formed from the Latin words “stolon,” and “ferum,” for “bearing.” While there is no completely standardized pronunciation for scientific names, the most common way to pronounce this species is probably so-LAY-num sto-lon-IF-er-um.
This species can survive frosts down to 27 degrees F (-3 C) (Li 1977). Vega (1995) found that this species is less frost tolerant than domesticated potato.
Although S. stoloniferum is tetraploid, it has disomic chromosome segregation, which probably explains why it is self-compatible.
Some accessions of this species appear to be edible and there was at least one farm that was growing S. cardiophyllum, S. ehrenbergii, and S. stoloniferum for market in Jalisco as recently as 2010 (Villa Vazquez 2010). Sotelo (1998) studied the nutrition of S. stoloniferum and found that it has a protein content about half to two-thirds that of the domesticated potato and vitamin C levels ranging from 2.93mg / 100g to 5.87 mg / 100g (as S. polytrichon).
This species is an allotetraploid, with two constituent genomes. Pendinen (2008) determined that the two genomes are derived from S. verrucosum and S. cardiophyllym, S. ehrenbergii, or S. jamesii.
Kiszonas (2010) measured the tuber pH of wild potato species and found that S. stoloniferum (as S. polytrichon) has a pH in the lower 10% of species tested. It is not known if tuber pH has any agronomic significance.
Solanum fendleri, a population found primarily in the United States, was recently absorbed into S. stoloniferum as a result of genetic analysis, but it appears that this grouping is somewhat controversial (Bamberg 2016).
Three accessions of this species, PI 660270, 667007, and 667008, have an unusual flower form, known as matryoshka for the nesting Russian dolls (Bamberg 2014). These flowers have extra whorls of petals and sometimes also extra whorls including anthers and stamens.
Subramanian (2017) found that at least some accessions of this species have unusually low potassium content (as S. fendleri).
In an analysis of 40 accessions of S. stoloniferum, Sanetomo (2021) found that 27 had D type cytoplasm and 13 had W type cytoplasm.
Resistances
Some accessions of this species carry the Rpi-sto1 and Rpi-pta1 genes, conferring late blight resistance.
S. stoloniferum has been the major source of extreme Potato Virus Y resistance in domesticated potato breeding.
| Condition | Type | Level of Resistance | Source |
|---|---|---|---|
| Alternaria solani (Early Blight) | Fungus | Somewhat resistant | Jansky 2008 |
| Drought | Abiotic | Somewhat resistant | Machida-Hirano 2015 (as S. fendleri) |
| Heat | Abiotic | Somewhat resistant | Machida-Hirano 2015 (as S. fendleri) |
| Globodera pallida (Pale Cyst Nematode) | Invertebrate | Somewhat resistant | Castelli 2003 (including S. fendleri) |
| Globodera pallida (Pale Cyst Nematode) | Invertebrate | Not resistant | Bachmann-Pfabe 2019 |
| Leptinotarsa decemlineata (Colorado Potato Beetle) | Invertebrate | Somewhat resistant | Machida-Hirano 2015 (as S. fendleri) |
| Meloidogyne spp. (Root Knot Nematode) | Invertebrate | Somewhat resistant | Machida-Hirano 2015 (as S. fendleri) |
| Myzus persicae (Green Peach Aphid) | Invertebrate | Resistant | Alvarez 2006, Alvarez (2013), Le Roux (2007) |
| Pectobacterium carotovorum (Blackleg/Soft Rot) | Bacteria | Not resistant | Chung 2011 |
| Globodera rostochiensis (Potato Cyst/Golden Nematode) | Invertebrate | Somewhat resistant | Castelli 2003 (including S. fendleri) |
| Phytophthora infestans (Late Blight) | Fungus | Resistant | Gonzales 2002, Bachmann-Pfabe 2019 |
| Phytophthora infestans (Late Blight) | Fungus | Not resistant | Gonzales 2002 (as S. fendleri) |
| Phytophthora infestans (Late Blight) | Fungus | Some resistance | Karki 2020 |
| Potato Leaf Roll Virus (PLRV) | Virus | Somewhat resistant | Machida-Hirano 2015 (as S. fendleri) |
| Potato Virus A (PVA) | Virus | Resistant | Wetter 1971 |
| Potato Virus Y (PVY) | Virus | Not resistant | Cai 2011 |
| Potato Virus Y (PVY) | Virus | Resistant | Wetter 1971 |
Glykoalkaloid content
Johns (1990) found glycoalkaloid levels of 64 and 69mg/100g for two accessions of this species (as S. fendleri). The primary glycoalkaloids were solanine and chaconine. Sotelo (1998) found TGA levels of 197mg/100g and 325mg/100g (as S. polytrichon). Despite these high levels, there are reports of S. stoloniferum (usually as S. fendleri) being eaten by native Americans and grown as an edible in Mexico. Most likely, there are either varieties with lower TGA levels or some sort of processing is done to lower glycoalkaloid content. Apparently, it was traditionally eaten with clay (White 1944, as S. fendleri) in order to eliminate some of the glycoalkaloids.
Images
 |
 |
 |
 |
 |
 |
 |
 |
Cultivation
Bamberg (2018) found that alternating temperatures increased germination in this species.
Towill (1983) found that seeds of this species stored at 1 to 3 degrees C germinated at 98 to 100% after 25 years. They also found that seeds of S. fendleri, now considered a synonym for this species, germinated at 28 to 92% after 21 years.
Breeding
Crosses with S. tuberosum
S. stoloniferum has been used often in potato breeding. Varieties that include S. stoloniferum in their pedigrees include Boone, Chenango, Hartford, Placid, Snowdrift (Hougas 1956), Conestoga, Pennchip, Pennrose (Plaisted 1989), and Bora Valley. The cultivar ‘Bora Valley’ has a 1/4 genetic contribution from S. stoloniferum (Choi 2003).
When used as the female in crosses with S. tuberosum, S. stoloniferum imposes tetrad sterility (T-CMS). Lines with a strain of the y mitochondrial genome (TSCsto), appear to always confer T-CMS, while those with normal cytoplasm are sometimes fertile (Sanetomo 2021).
The expected result of a cross of S. stoloniferum x S. tuberosum (4x) would be a hexaploid formed by the union of an unreduced egg from S. stoloniferum and normal haploid pollen from S. tuberosum, resulting in a zygote with four copies of the S. stoloniferum genome and two copies of the S. tuberosum genome. Interestingly, other configurations have been produced. Panahandeh (2019) describes obtaining several tetraploid S. stoloniferum x 4x S. tuberosum hybrids through conventional crossing, a combination that has a low probability due to EBN incompatibility. Analysis of the chromosome pairing suggested that the genome composition of the progeny was AAAB. Sanetomo (2021) noted a number of tetraploid and triploid progeny that were determined be true crosses and proposed double reduction in the S. tuberosum pollen or genome elimination as possible causes.
| Female | Male | Berry Set |
Seed Set | Ploidy | Germ | Source |
|---|---|---|---|---|---|---|
| S. tuberosum | S. stoloniferum (as S. fendleri and S. stoloniferum) | Low | None | Jackson (1999) | ||
| S. tuberosum | S. stoloniferum (as S. papita) | Low | High | Jackson (1999) | ||
| S. stoloniferum (as S. fendleri and S. stoloniferum) | S. tuberosum | Low | Low | Jackson (1999) | ||
| S. stoloniferum |
S. tuberosum (4x) |
Moderate | Low | 6x, 4x, 3x | Yes | Sanetomo (2021) |
Crosses with other species
Watanabe (1991) found that 8.7% of varieties of this species produced 2n pollen (calculated from the totals for S. stoloniferum, S. fendleri, S. papita, and S. polytrichon, which have been reclassified as S. stoloniferum) which would be effectively octaploid and 4EBN. Jackson (1999) found 0-29%.
| Female | Male | Berry Set |
Seed Set | Ploidy | Germ | Source |
|---|---|---|---|---|---|---|
| S. commersonii 3x | S. stoloniferum |
Yes | Yes | Yes | Vavilov 1938 | |
| S. commersonii 3x | S. stoloniferum | Yes | Yes | Yes | Emme 1937 | |
| S. commersonii |
S. stoloniferum (as S. fendleri) | Yes | Yes | Reddick 1939 | ||
| S. stoloniferum | 4x 2EBN S. cardiophyllum | None | None | Hayes (2005) | ||
| S. stoloniferum (as S. fendleri) | S. commersonii | Yes | Yes | Reddick 1939 | ||
| S. stoloniferum | 4x 2EBN S. pinnatisectum | None | None | Hayes (2005) | ||
| 4x 2EBN S. pinnatisectum | S. stoloniferum | None | None | Hayes (2005) |
References
Solanum stoloniferum at Solanaceae Source